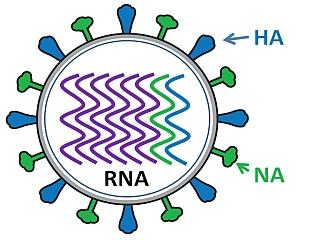
Influenza virus
Influenza virus has a rounded shape (although it can be elongated or irregularly shaped) and has a layer of spikes on the outside. There are two different kinds of spikes, each made of a different protein – one is the hemagglutinin (HA) protein and the other is the neuraminidase (NA) protein. Inside the layer of spikes, there are eight single-stranded viral RNA contained in separate viral ribonucleoprotein (vRNP) complexes.
The influenza A, B, and C viruses, representing three of the five genera of the family Orthomyxoviridae, are characterized by segmented, negative-strand RNA genomes, requiring an RNA-dependent RNA polymerase of viral origin for replication. The particular structure of the influenza virus genome and function of its viral proteins enable antigenic drift and antigenic shift. These processes result in viruses able to evade the long-term adaptive immune responses in many hosts.
Matrix protein 2 (Q76V12)
Function
Forms a proton-selective ion channel that is necessary for the efficient release of the viral genome during virus entry. After attaching to the cell surface, the virion enters the cell by endocytosis. Acidification of the endosome triggers M2 ion channel activity. The influx of protons into virion interior is believed to disrupt interactions between the viral ribonucleoprotein (RNP), matrix protein 1 (M1), and lipid bilayers, thereby freeing the viral genome from interaction with viral proteins and enabling RNA segments to migrate to the host cell nucleus, where influenza virus RNA transcription and replication occur. Also plays a role in viral proteins secretory pathway. Elevates the intravesicular pH of normally acidic compartments, such as trans-Golgi network, preventing newly formed hemagglutinin from premature switching to the fusion-active conformation.
Matrix protein 1 (Q67157)
Function
Plays critical roles in virus replication, from virus entry and uncoating to assembly and budding of the virus particle. M1 binding to ribonucleocapsids (RNPs) in nucleus seems to inhibit viral transcription. Interaction of viral NEP with M1-RNP is thought to promote nuclear export of the complex, which is targeted to the virion assembly site at the apical plasma membrane in polarized epithelial cells. Interactions with NA and HA may bring M1, a non-raft-associated protein, into lipid rafts. Forms a continuous shell on the inner side of the lipid bilayer in virion, where it binds the RNP. During virus entry into cell, the M2 ion channel acidifies the internal virion core, inducing M1 dissociation from the RNP. M1-free RNPs are transported to the nucleus, where viral transcription and replication can take place.
Hemagglutinin (P03437)
Function
Binds to sialic acid-containing receptors on the cell surface, bringing about the attachment of the virus particle to the cell. This attachment induces virion internalization of about two third of the virus particles through clathrin-dependent endocytosis and about one third through a clathrin- and caveolin-independent pathway. Plays a major role in the determination of host range restriction and virulence. Class I viral fusion protein. Responsible for penetration of the virus into the cell cytoplasm by mediating the fusion of the membrane of the endocytosed virus particle with the endosomal membrane. Low pH in endosomes induces an irreversible conformational change in HA2, releasing the fusion hydrophobic peptide. Several trimers are required to form a competent fusion pore.
Binds to sialic acid-containing receptors on the cell surface, bringing about the attachment of the virus particle to the cell. This attachment induces virion internalization either through clathrin-dependent endocytosis or through clathrin- and caveolin-independent pathway. Plays a major role in the determination of host range restriction and virulence. Class I viral fusion protein. Responsible for penetration of the virus into the cell cytoplasm by mediating the fusion of the membrane of the endocytosed virus particle with the endosomal membrane. Low pH in endosomes induces an irreversible conformational change in HA2, releasing the fusion hydrophobic peptide. Several trimers are required to form a competent fusion pore.
Nuclear export protein (Q02600)
Function
Mediates the nuclear export of encapsidated genomic RNAs (ribonucleoproteins, RNPs). Acts as an adapter between viral RNPs complexes and the nuclear export machinery of the cell. Possesses no intrinsic RNA-binding activity, but includes a C-terminal M1-binding domain. This domain is believed to allow recognition of RNPs bound to the protein M1. Since protein M1 is not available in large quantities before late stages of infection, such an indirect recognition mechanism probably ensures that genomic RNPs are not exported from the host nucleus until sufficient quantities of viral mRNA and progeny genomic RNA have been synthesized. Furthermore, the RNPs enter the host cytoplasm only when associated with the M1 protein that is necessary to guide them to the plasma membrane. May down-regulate viral RNA synthesis when overproduced.
Non-structural protein 1 (P69277)
Function
Prevents the establishment of the cellular antiviral state by inhibiting TRIM25-mediated DDX58 ubiquitination, which normally triggers the antiviral transduction signal that leads to the activation of type I IFN genes by transcription factors IRF3 and IRF7. Prevents human EIF2AK2/PKR activation, either by binding double-strand RNA, or by interacting directly with EIF2AK2/PKR. This function may be important at the very beginning of the infection, when NS1 is mainly present in the cytoplasm. Also binds poly(A) and U6 snRNA.
Inhibits post-transcriptional processing of cellular pre-mRNA, by binding and inhibiting two cellular proteins that are required for the 3'-end processing of cellular pre-mRNAs: the 30 kDa cleavage and polyadenylation specificity factor/CPSF4 and the poly(A)-binding protein 2/PABPN1. In turn, unprocessed 3' end pre-mRNAs accumulate in the host nucleus and are no longer exported to the cytoplasm. Cellular protein synthesis is thereby shut off very early after virus infection. Viral protein synthesis is not affected by the inhibition of the cellular 3' end processing machinery because the poly(A) tails of viral mRNAs are produced by the viral polymerase through a stuttering mechanism.
Neuraminidase (Q75VQ4)
Function
Catalyzes the removal of terminal sialic acid residues from viral and cellular glycoconjugates. Cleaves off the terminal sialic acids on the glycosylated HA during virus budding to facilitate virus release. Additionally helps virus spread through the circulation by further removing sialic acids from the cell surface. These cleavages prevent self-aggregation and ensure the efficient spread of the progeny virus from cell to cell. Otherwise, infection would be limited to one round of replication. Described as a receptor-destroying enzyme because it cleaves a terminal sialic acid from the cellular receptors. May facilitate viral invasion of the upper airways by cleaving the sialic acid moieties on the mucin of the airway epithelial cells. Likely to plays a role in the budding process through its association with lipid rafts during intracellular transport. May additionally display a raft-association independent effect on budding. Plays a role in the determination of host range restriction on replication and virulence. Sialidase activity in late endosome/lysosome traffic seems to enhance virus replication.
Reference:
1.Bouvier NM, Palese P. The biology of influenza viruses. Vaccine. 2008;26 Suppl 4(Suppl 4):D49-D53. doi:10.1016/j.vaccine.2008.07.039
2.Dadonaite B, Gilbertson B, Knight ML, Trifkovic S, Rockman S, Laederach A, Brown LE, Fodor E, Bauer DLV. The structure of the influenza A virus genome. Nat Microbiol. 2019 Nov;4(11):1781-1789. doi: 10.1038/s41564-019-0513-7. Epub 2019 Jul 22. PMID: 31332385; PMCID: PMC7191640.
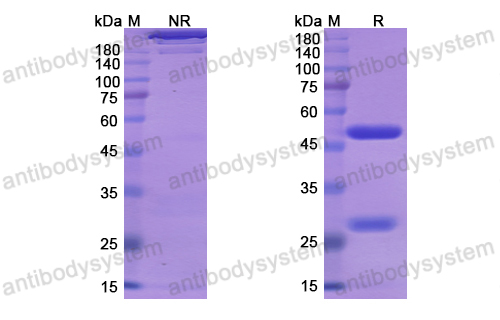
Host species: Human
Isotype: IgG1, kappa
Applications: Research Grade Biosimilar
Expression system: Mammalian Cells
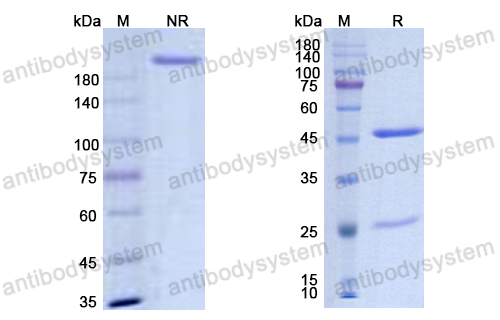
Host species: Human
Isotype: IgG1, lambda
Applications: Research Grade Biosimilar
Expression system: Mammalian Cells
Host species: Human
Isotype: IgG1, lambda
Applications: ELISA, Neutralization

Host species: Human
Isotype: IgG1, kappa
Applications: ELISA, Neutralization
Host species: Human
Isotype: IgG1, kappa
Applications: ELISA, Neutralization
Accession: M1UPL4
Applications: ELISA, Immunogen, SDS-PAGE, WB, Bioactivity testing in progress
Expression system: Mammalian Cells
Accession: A0A248XKH7
Protein length: Gln17-Tyr527
Host species: Human
Isotype: IgG1
Applications: ELISA, Neutralization
Accession: A0A076G8X7
Host species: Human
Isotype: IgG1
Applications: ELISA, Neutralization
Accession: Q6DQ34
Host species: Human
Isotype: IgG1
Applications: ELISA, Neutralization
Accession: Q67333
Applications: ELISA, Immunogen, SDS-PAGE, WB, Bioactivity testing in progress
Expression system: E. coli
Accession: P03430
Protein length: Val286-Tyr483



