Syphilis is a complex sexually transmitted, chronic and multistage disease caused by Treponema pallidum, affects approximately 36 million people globally, with 11 million new infections each year. The first T. pallidum strain was isolated in 1912.
T. pallidum, previously referred to as Spirochaeta pallida, is a microaerophilic spirochete bacterium responsible for causing diseases such as syphilis, endemic syphilis (also known as bejel), and yaws. This bacterium is known to infect only humans and baboons. T. pallidum is a helically coiled organism, typically measuring between 6-15 μm in length and 0.1-0.2 μm in width. Due to the absence of a tricarboxylic acid cycle and oxidative phosphorylation, it exhibits very limited metabolic activity. The Treponemes possess both cytoplasmic and outer membranes and can only be observed under light microscopy with dark-field illumination.
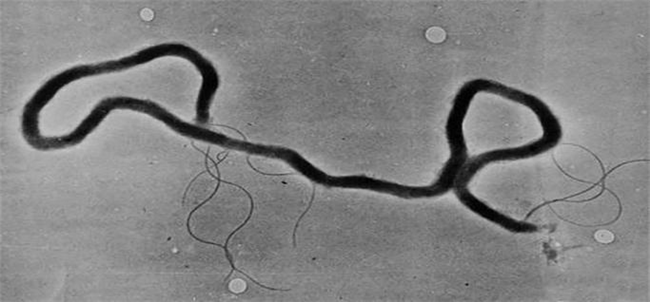
Figure 1. Electron micrograph of Treponema pallidum
T. pallidum includes three subspecies: T. p. pallidum, T. p. endemicum, and T. p. pertenue, each associated with different diseases. The bacterium is helically shaped and highly motile, comprising an outer membrane, a peptidoglycan layer, an inner membrane, a protoplasmic cylinder, and a periplasmic space. Although often classified as Gram-negative, its outer membrane lacks the lipopolysaccharides typical of other Gram-negative bacteria. Instead, it features an endoflagellum located within the periplasmic space, wrapping around the protoplasmic cylinder, and consisting of four main polypeptides, a core structure, and a sheath.
The global public health threat posed by syphilis underscores the need for effective control measures. Despite the availability of inexpensive and effective penicillin treatment for over 70 years, the continued high incidence of syphilis worldwide highlights the urgent need for alternative approaches to syphilis control.
The identification of T. pallidum proteins began in 1975 with the application of electrophoretic techniques. The outer membrane of T. pallidum interacts directly with host cells and contains very few transmembrane proteins, which limits its antigenicity. It is covered in lipoproteins that facilitate its attachment to host cells through specific treponemal ligands, which are functionally and antigenically similar. Additionally, the genus Treponema has ribbons of cytoskeletal filaments, similar to intermediate filaments, running along the length of the cell just beneath the cytoplasmic membrane, providing structural support.
The proteome of T. pallidum consists of 998 proteins. Some of these proteins are used for the diagnostic detection of T. pallidum, while others are considered as vaccine candidates.
Tpn15/TP_0171(P16055)
The function of Tpn15 remains unknown, but it is hypothesized to be a flavoprotein. The study found that Tpn15 is strongly expressed across different stages of syphilis.
Tpn17/TP_0435(P29722)
Tpn17 is a 17-kDa lipoprotein and a well konwn syphilis antigen. Despite making up only about 2% of the detergent-soluble protein complement of T. pallidum, Tpn17 is a potent immunogen. It induces proliferation in syphilitic rabbit splenocytes and stimulates the production of TNF-α, an inflammation signaling factor, by murine macrophages. The Tpn17 gene is also one of the most highly expressed genes in T. pallidum during rabbit infection. Previous studies have shown that Tpn17 can oligomerize under oxidizing conditions but reverts to monomers when treated with reducing agents. Despite this wealth of information, the exact function of Tpn17 remains unclear.
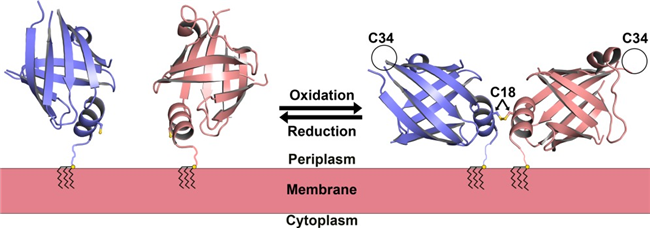
Figure 2. The modeled in vivo topology of the TP0435 dimer (Brautigam et al., 2014)
Tpn47/TP_0574(P29723)
Tpn47 is a 47-kDa lipoprotein hypothesized to be a zinc dependent D,D-carboxypeptidase and functions as a penicillin-binding protein (PBP). Tpn47 can binds penicillin covalently. It contains three PBP signature motifs (SVTK, TEN, and KTG), but they do not form a typical active site. The crystal structure reveals a unique fold, predominantly composed of beta-sheets, with four distinct domains: two beta-sheet-rich N-terminal domains and two C-terminal domains with immunoglobulin-like folds, differing significantly from other known PBPs.
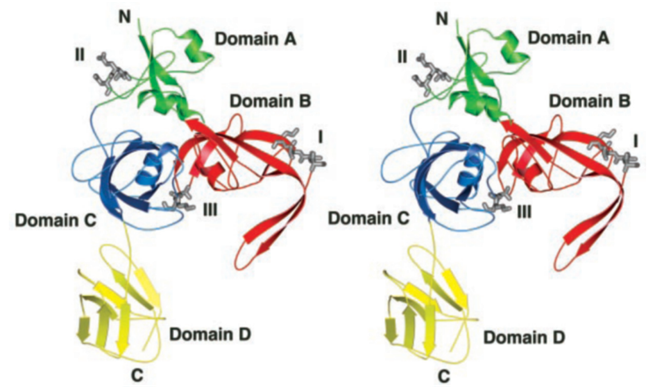
Figure 3 Stereoview of the T. pallidum Tpn47 ((Deka et al., 2002)
In a phase I study, TpN15, TpN17, TpN47 and TmpA recombinant proteins exhibited superior diagnostic performance. TpN17 and TmpA showed the most promising results and advanced to a phase II study. Several other studies have explored these proteins in the development of serological tests, seeking precision, rapidity, and cost-effectiveness
Tp0954
Animals immunized with recombinant Tp0954 (rTp0954) produced high titers of Tp0954-specific serum IgG and showed elevated levels of IFN-γ from splenocytes, along with a specific splenocyte proliferation response. In comparison, control animals immunized with PBS and Freund’s adjuvant (FA) did not exhibit these responses. Furthermore, rTp0954 immunization significantly delayed the development of cutaneous lesions, promoted inflammatory cellular infiltration at the primary lesion sites, and inhibited the dissemination of *T. pallidum* to distal tissues or organs.
Tp0751 (O83732)
Tp0751 possesses adhesive properties and has been shown to mediate attachment to host extracellular matrix components such as laminin, fibronectin, and fibrinogen. In studies, Tp0751-immunized animals exhibited a significantly reduced bacterial burden in organs after T. pallidum challenge compared to unimmunized animals. Furthermore, transferring lymph nodes from Tp0751-immunized and T. pallidum-challenged animals to naïve animals did not induce infection, confirming sterile protection. These findings suggest that Tp0751 is a promising candidate for a syphilis vaccine.
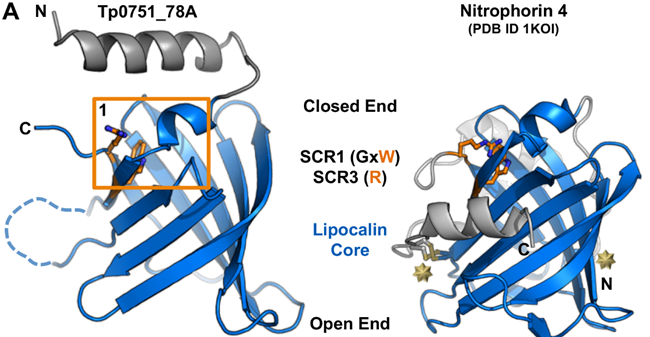
Figure 4. Structure of Tp0751
TprK
Immunization with full-length rTprK was found to be effective in attenuating lesion development and accelerating lesion resolution, thereby reducing the likelihood of the pathogen spreading to distant tissues and potentially preventing disease progression. Additionally, the results showed that rTprK immunization rapidly induced a strong Th1-like cellular response and elicited a humoral immune response, producing opsonic antibodies that enhanced macrophage-mediated opsonophagocytosis. While complete protection against infection was not achieved, the study provided a thorough exploration of the immunogenicity of TprK and underscored its potential as a key component in a syphilis vaccine.
Tp0435(O67998)
TP0453 is a unique lipoprotein in the outer membrane of Treponema pallidum, capable of integrating into membranes through its amphipathic helices, with two adjacent helices critical for sensing and insertion into the lipid bilayer. It exhibits dynamic structural transitions between closed and open conformations, exposing a hydrophobic cavity that likely facilitates lipid interactions. TP0453 also enhances membrane permeability under acidic conditions, suggesting a porin-like activity. Additionally, its structural similarity to certain Mycobacterium tuberculosis lipoproteins suggests it functions as a carrier of lipids or glycolipids during outer membrane biogenesis in T. pallidum.
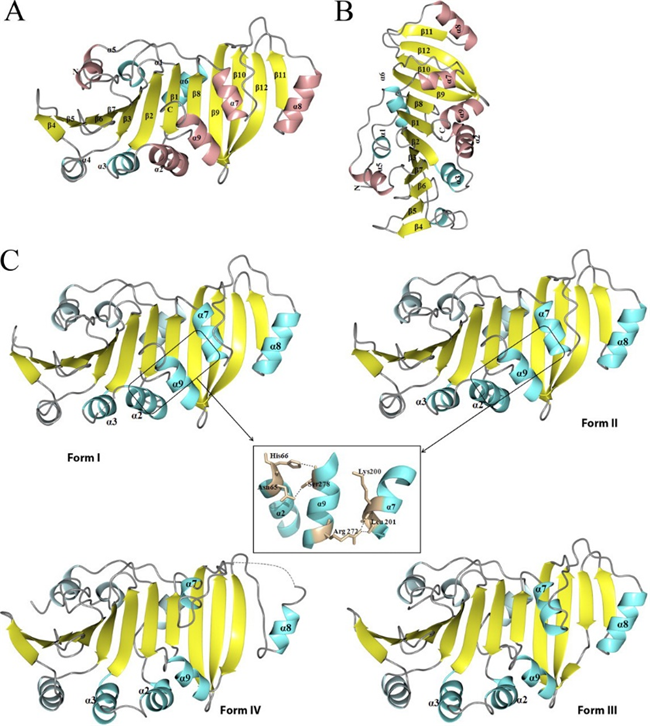
Figure 5. Crystal structure of Tp0435
Reference
1. Baughn, R.E., Demecs, M., Taber, L.H., and Musher, D.M. (1996). Epitope mapping of B-cell determinants on the 15-kilodalton lipoprotein of Treponema pallidum (Tpp15) with synthetic peptides. Infect Immun 64, 2457-2466.
2. Brautigam, C.A., Deka, R.K., Liu, W.Z., and Norgard, M.V. (2014). Insights into the potential function and membrane organization of the TP0435 (Tp17) lipoprotein from Treponema pallidum derived from structural and biophysical analyses. Protein Science 24, 11-19.
3. Cameron, C.E. (2018). Syphilis Vaccine Development: Requirements, Challenges, and Opportunities. Sex Transm Dis 45, S17-S19.
4. Deka, R.K., Machius, M., Norgard, M.V., and Tomchick, D.R. (2002). Crystal structure of the 47-kDa lipoprotein of Treponema pallidum reveals a novel penicillin-binding protein. Journal of Biological Chemistry 277, 41857-41864.
5. Gomes, L.G.R., Rodrigues, T.C.V., Jaiswal, A.K., Santos, R.G., Kato, R.B., Barh, D., Alzahrani, K.J., Banjer, H.J., Soares, S.d.C., and Azevedo, V. (2022). In Silico Designed Multi-Epitope Immunogen “Tpme-VAC/LGCM-2022” May Induce Both Cellular and Humoral Immunity against Treponema pallidum Infection. Vaccines 10, 1019.
6. He, Y., Chen, D., Fu, Y., Huo, X., Zhao, F., Yao, L., Zhou, X., Qi, P., Yin, H., Cao, L., et al. (2023). Immunization with Tp0954, an adhesin of Treponema pallidum, provides protective efficacy in the rabbit model of experimental syphilis. Front Immunol 14, 1130593.
7. Kojima, N., Konda, K.A., and Klausner, J.D. (2022). Notes on syphilis vaccine development. Front Immunol 13, 952284.
8. Kubanov, A., Runina, A., and Deryabin, D. (2017). Novel Treponema pallidum Recombinant Antigens for Syphilis Diagnostics: Current Status and Future Prospects. Biomed Res Int 2017, 1436080.
9. Kumar Jaiswal, A., Tiwari, S., Jamal, S.B., Barh, D., Azevedo, V., and Soares, S.C. (2017). An In Silico Identification of Common Putative Vaccine Candidates against Treponema pallidum: A Reverse Vaccinology and Subtractive Genomics Based Approach. Int J Mol Sci 18, 402.
10. Lithgow, K.V., Hof, R., Wetherell, C., Phillips, D., Houston, S., and Cameron, C.E. (2017). A defined syphilis vaccine candidate inhibits dissemination of Treponema pallidum subspecies pallidum. Nat Commun 8, 14273.
11. Liu, D., Chen, R., Wang, Y.-J., Li, W., Liu, L.-L., Lin, L.-R., Yang, T.-C., and Tong, M.-L. (2023). Insights into the protective immune response by immunization with full-length recombinant TprK protein: cellular and humoral responses. npj Vaccines 8, 146.
12. Luthra, A., Zhu, G., Desrosiers, D.C., Eggers, C.H., Mulay, V., Anand, A., McArthur, F.A., Romano, F.B., Caimano, M.J., Heuck, A.P., et al. (2011). The Transition from Closed to Open Conformation of Treponema pallidum Outer Membrane-associated Lipoprotein TP0453 Involves Membrane Sensing and Integration by Two Amphipathic Helices *. Journal of Biological Chemistry 286, 41656-41668.
13. Morshed, M.G., and Singh, A.E. (2015). Recent trends in the serologic diagnosis of syphilis. Clin Vaccine Immunol 22, 137-147.
14. Osias, E., Hung, P., Giacani, L., Stafylis, C., Konda, K.A., Vargas, S.K., Reyes-Diaz, E.M., Comulada, W.S., Haake, D.A., Haynes, A.M., et al. (2020). Investigation of syphilis immunology and Treponema pallidum subsp. pallidum biology to improve clinical management and design a broadly protective vaccine: study protocol. BMC Infect Dis 20, 444.
15. Parker, M.L., Houston, S., Pětrošová, H., Lithgow, K.V., Hof, R., Wetherell, C., Kao, W.-C., Lin, Y.-P., Moriarty, T.J., and Ebady, R. (2016). The structure of Treponema pallidum Tp0751 (Pallilysin) reveals a non-canonical lipocalin fold that mediates adhesion to extracellular matrix components and interactions with host cells. PLoS Pathogens 12, e1005919.
16. Silva, A.A.O., Lima, A.A., Vasconcelos, L.C.M., Almeida, R.A., Freitas, N.E.M., Oliva, T.A., de Carvalho Ribeiro da Silva, M.F., Marchini, F.K., Zanchin, N.I.T., de Siqueira, I.C., et al. (2024). Evaluating the diagnostic accuracy of TpN17 and TmpA recombinant proteins in syphilis detection: a phase II study. Front Microbiol 15, 1348437.
17. Sun, A.H., Mao, Y.F., Hu, Y., Sun, Q., and Yan, J. (2009). Sensitive and specific ELISA coated by TpN15-TpN17-TpN47 fusion protein for detection of antibodies to Treponema pallidum. Clin Chem Lab Med 47, 321-326.
18. Xu, M., Xie, Y., Zheng, K., Luo, H., Tan, M., Zhao, F., Zeng, T., and Wu, Y. (2021). Two Potential Syphilis Vaccine Candidates Inhibit Dissemination of Treponema pallidum. Front Immunol 12, 759474.
19. Young, H., Moyes, A., Seagar, L., and McMillan, A. (1998). Novel recombinant-antigen enzyme immunoassay for serological diagnosis of syphilis. J Clin Microbiol 36, 913-917.

Host species: Rabbit
Isotype: IgG
Applications: ELISA, IHC, WB
Accession: P16665
Applications: ELISA, Immunogen, SDS-PAGE, WB, Bioactivity testing in progress
Expression system: E. coli
Accession: P07643
Protein length: Ser21-Arg345
Applications: ELISA, Immunogen, SDS-PAGE, WB, Bioactivity testing in progress
Expression system: E. coli
Accession: P29723
Protein length: Cys20-Gln434
Applications: ELISA, Immunogen, SDS-PAGE, WB, Bioactivity testing in progress
Expression system: E. coli
Accession: P16055 + WP_010882213.1 + WP_010882021.1
Protein length: TPP15 [Ser19-Arg141] + tmpA [Gly20-Arg345] + TPP47 [Gly21-Gln434]

Applications: ELISA, Immunogen, SDS-PAGE, WB, Bioactivity testing in progress
Expression system: E. coli
Accession: P29722
Protein length: Cys22-Lys156
Applications: ELISA, Immunogen, SDS-PAGE, WB, Bioactivity testing in progress
Expression system: E. coli
Accession: P16665
Protein length: Met1-Ala177




