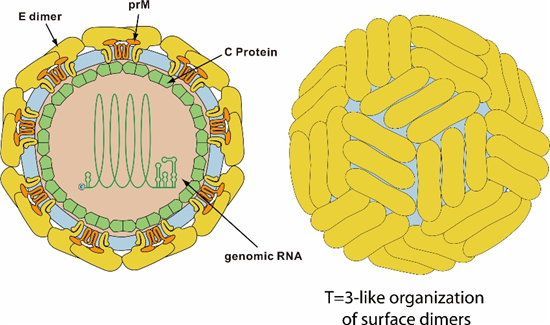
Fig. 1 Structure of Japanese Encephalitis Virus (ViralZone)
Japanese Encephalitis (JE) is a severe neurological disease caused by infection with the Japanese Encephalitis Virus (JEV). It is the most important viral encephalitis worldwide, with approximately 35,000 to 50,000 cases reported annually. The mortality rate is estimated to be between 10,000 and 15,000 deaths per year.
JEV is a mosquito-borne arbovirus, part of the Flaviviridae family, which also includes other notable viruses such as yellow fever virus, West Nile virus (WNV), dengue virus (DENV), and Zika virus (ZIKV). The first documented case of JEV infection occurred in 1871 in Japan, and the disease was later named Japanese Encephalitis (JE).
JEV is a single-stranded, positive-sense RNA virus. Its genome is approximately 11 kb in length and contains a single open reading frame (ORF) that encodes a long polyprotein. This polyprotein is co- and post-translationally processed by a combination of viral and cellular proteases into three structural proteins: the capsid protein (C), the non-glycosylated pre-membrane protein (prM), and the glycosylated envelope protein (E). In addition, the virus produces seven non-structural (NS) proteins: NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5.
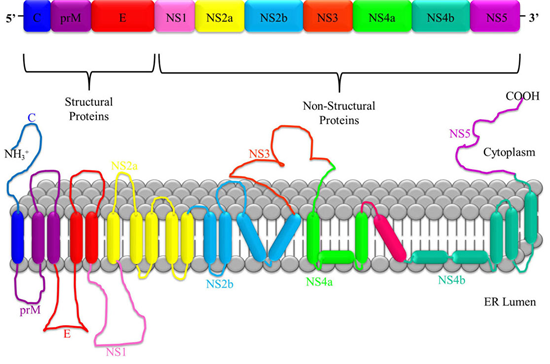
Fig. 2 The genome and polypeptide structure of Japanese Encephalitis Virus (Roberts and Gandhi, 2020)
Capsid Protein
The nucleocapsid of JEV consists of multiple copies of the capsid protein, which encloses the single-stranded RNA genome. The capsid protein forms an inner shell and is essential for the viral lifecycle, displaying a helix-rich structure and forming a stable homodimer, similar to other flaviviruses. The N-terminus of the capsid is unstable, and α-helix-1 is flexible, adopting a closed conformation in JEV. The α4-4’ site on the dimeric interface may interact with viral RNA due to its coiled-coil-like structure.
The monomeric capsid protein shares structural similarities with nucleic acid-binding proteins. It plays a crucial role in virus budding by binding to the cell membrane and packaging viral RNA into the nucleocapsid. During virus entry, it may aid in genome penetration after hemifusion induced by surface proteins. Additionally, the capsid can migrate to the cell nucleus, modulating host functions, and it counteracts antiviral responses by sequestering and degrading host EXOC1 via the proteasome degradation pathway.
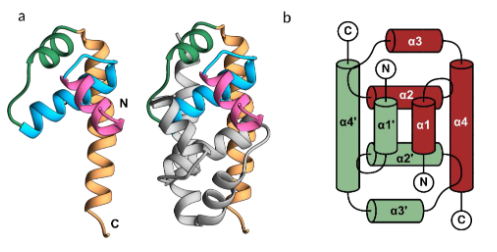
Fig. 3 The structure of the Japanese encephalitis virus (JEV) capsid protein (Poonsiri et al., 2019)
prM
PrM is an outer membrane protein with lipid-binding and metal-binding properties, belonging to the antibiotic resistance protein family, which helps nullify the action of antibiotics. It prevents premature fusion of the envelope proteins in the trans-Golgi network by binding to envelope protein E at pH 6.0. After the virion is released into the extracellular space, PrM dissociates from the E dimers, allowing the virion to mature and become fully infectious.
Envelope protein E
The E protein of the Japanese Encephalitis Virus (JEV) mediates host-virus attachment and fusion between the viral and host cell membranes. The E protein is a homodimer consisting of three structural domains: D1, D2, and D3, with domain D3 (ED3) being the primary site for neutralizing antibodies. Most of these neutralizing agents target the terminal end of ED3. It is a promising vaccine candidate.

Fig. 4 Crystal structure of JEV E ectodomain (Luca et al., 2012)
NS1
NS1 is a highly conserved non-structural protein found in flaviviruses, existing in both intracellular dimer and secreted hexamer forms. The dimer plays a role in genome replication, while the hexamer contributes to immune evasion. Secreted NS1 has been identified as a potential diagnostic marker for early detection of flavivirus infections and is also being explored for therapeutic development, with NS1-based subunit vaccines in various stages of development. Once cleaved from the polyprotein, NS1 is targeted to three key locations: the viral replication cycle, the plasma membrane, and the extracellular compartment, where it is essential for viral replication, pathogenesis, and immune evasion.
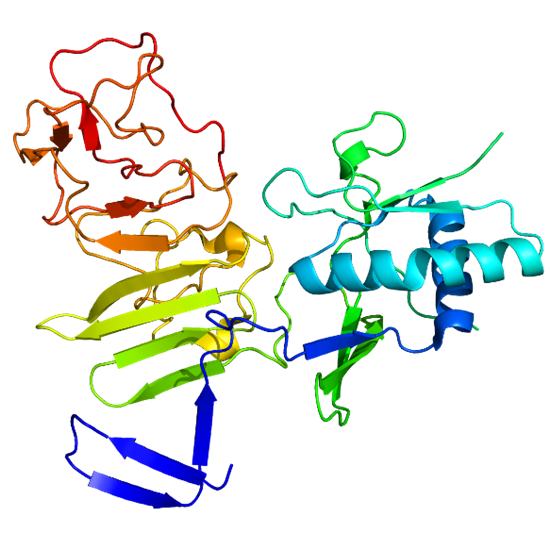
Fig. 5 Crystal structure of NS1 (Pan et al., 2024)
Reference
1. Luca, V.C., AbiMansour, J., Nelson, C.A., and Fremont, D.H. (2012). Crystal Structure of the Japanese Encephalitis Virus Envelope Protein. Journal of Virology 86, 2337-2346.
2. Pan, Q., Jiao, H., Zhang, W., Chen, Q., Zhang, G., Yu, J., Zhao, W., and Hu, H. (2024). The step-by-step assembly mechanism of secreted flavivirus NS1 tetramer and hexamer captured at atomic resolution. Science Advances 10, eadm8275.
3. Poonsiri, T., Wright, G.S.A., Solomon, T., and Antonyuk, S.V. (2019). Crystal Structure of the Japanese Encephalitis Virus Capsid Protein. Viruses 11, 623.
4. Rastogi, M., Sharma, N., and Singh, S.K. (2016). Flavivirus NS1: a multifaceted enigmatic viral protein. Virology Journal 13.
5. Roberts, A., and Gandhi, S. (2020). Japanese encephalitis virus: a review on emerging diagnostic techniques. Front Biosci (Landmark Ed) 25, 1875-1893.
6. Roy, U. (2020). Structural and molecular analyses of functional epitopes and escape mutants in Japanese encephalitis virus envelope protein domain III. Immunol Res 68, 81-89.
7. Srivastava, K.S., Jeswani, V., Pal, N., Bohra, B., Vishwakarma, V., Bapat, A.A., Patnaik, Y.P., Khanna, N., and Shukla, R. (2023). Japanese Encephalitis Virus: An Update on the Potential Antivirals and Vaccines. Vaccines 11, 742.
8. Xiong, J., Yan, M., Zhu, S., Zheng, B., Wei, N., Yang, L., Si, Y., Cao, S., and Ye, J. (2022). Increased Cleavage of Japanese Encephalitis Virus prM Protein Promotes Viral Replication but Attenuates Virulence. Microbiol Spectr 10, e0141722.
Host species: Mouse
Isotype: IgG2a, kappa
Applications: ELISA, Neutralization
Accession: P14403

Host species: Mouse
Isotype: IgG2a, kappa
Applications: ELISA, Neutralization
Accession: P14403
Host species: Mouse
Isotype: IgG2a, kappa
Applications: ELISA, Neutralization
Accession: P14403
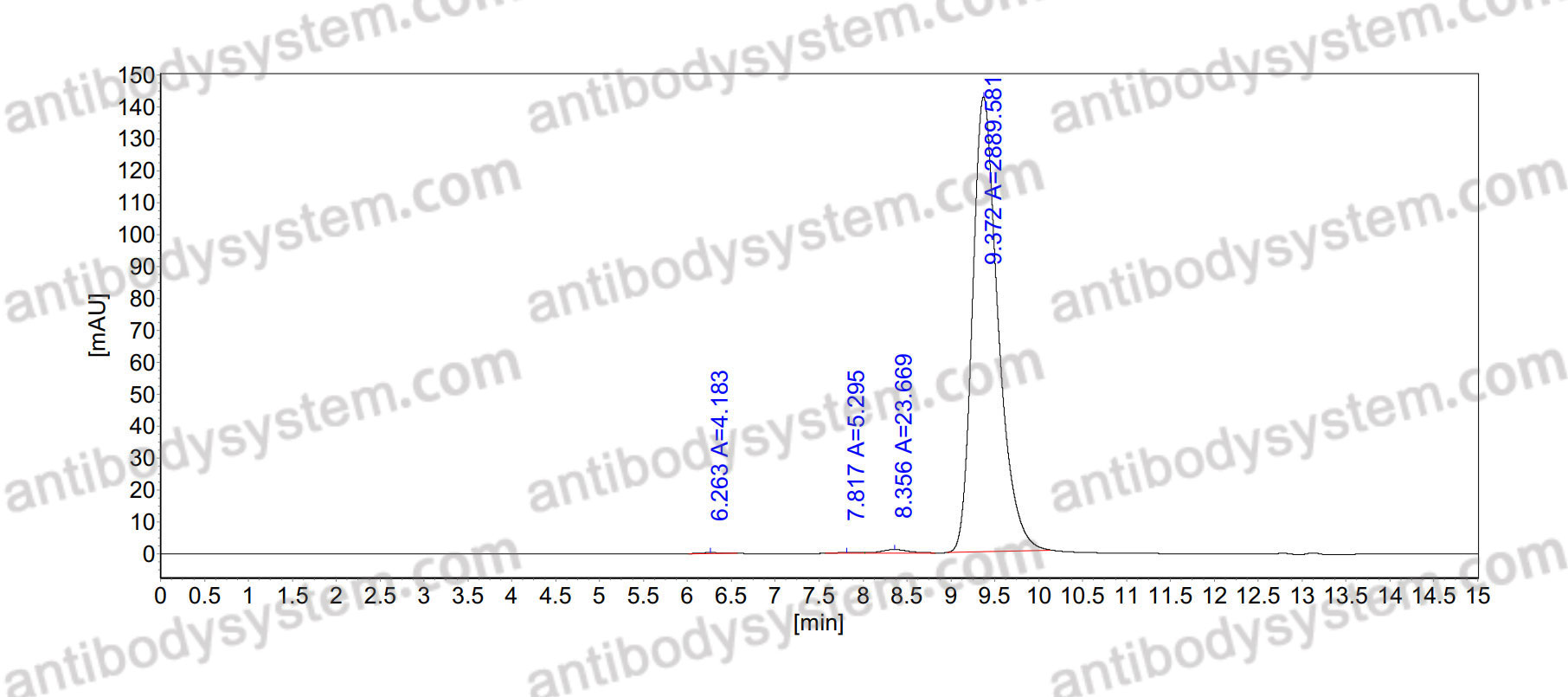
Host species: Rabbit
Isotype: IgG
Applications: ELISA, IHC, WB
Accession: P0DOK8
Host species: Rabbit
Isotype: IgG
Applications: ELISA, IHC, WB
Accession: P0DOK8
Host species: Rabbit
Isotype: IgG
Applications: ELISA, IHC, WB
Accession: NP_059434.1
Host species: Rabbit
Isotype: IgG
Applications: ELISA, IHC, WB
Accession: P19110
Host species: Rabbit
Isotype: IgG
Applications: ELISA, IHC, WB
Accession: P32886
Host species: Rabbit
Isotype: IgG
Applications: ELISA, IHC, WB
Accession: P09732
Host species: Rabbit
Isotype: IgG
Applications: ELISA, IHC, WB
Accession: P09732



