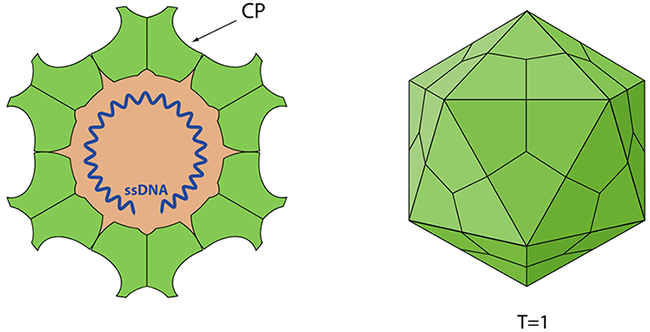
Figure 1 Schematic representation of Canine Parvovirus (viralzone)
Canine Parvovirus Type 2 (CPV-2) is a highly contagious viral disease that predominantly affects domestic dogs. Identified in the late 1970s, CPV-2 has since emerged as a critical health concern for dogs worldwide. The virus is notorious for its rapid spread and severe clinical manifestations, leading to high mortality rates, especially in puppies.
CPV-2 is believed to have originated as a host range variant from feline panleucopenia virus (FPV), include a direct mutation from FPV, a mutation from a FPV vaccine virus and the adaptation to the new host dog via non-domestic carnivores, like mink and foxes. The virus has several variants, distinguished by their surface protein (VP capsid) characteristics. The first identified variants, CPV-2a and CPV-2b, emerged in 1979 and 1984, respectively. A more recent variant, CPV-2c, which involves a Glu-426 mutation, has been discovered in countries including Italy, Vietnam, and Spain.
CPV-2 is an exceptionally small virus, measuring only 20 to 26 nanometers in diameter, with an icosahedral symmetry. As a member of the Parvoviridae family, CPV-2 is a non-enveloped, single-stranded DNA virus. Parvoviruses are known for their ability to infect both vertebrates and insects. This structural simplicity contributes to the virus's resilience, enabling it to survive in harsh environmental conditions and making eradication challenging.
Given the virulent and highly contagious nature of CPV-2, prevention is the most effective strategy to ensure the health of puppies and adult dogs. It is essential to begin vaccination when puppies are 7 to 8 weeks old, with booster shots administered every 3 to 4 weeks until they are at least 16 weeks old. While the antigenic patterns of CPV-2a and CPV-2b closely resemble the original CPV-2, the CPV-2c variant has a unique antigenicity. However, studies have shown that existing vaccines based on CPV-2b provide sufficient protection against CPV-2c as well.
Capsid protein VP1 (P12930)
The canine parvovirus (CPV) capsid is described as having a seemingly simple structure. This capsid is constructed from 60 identical subunits composed of two distinct proteins: virus protein 1 (VP1), which has a molecular weight of 84 kDa, and virus protein 2 (VP2), weighing 67 kDa. VP2 constitutes about 90% of the capsid's protein content, while VP1 makes up the remaining 10%. Since VP2 in the CPV capside is the most abundant protein. It is reasonable to think that VP2 could form virus like particles (VLPs). It is a promising vaccine candidate.
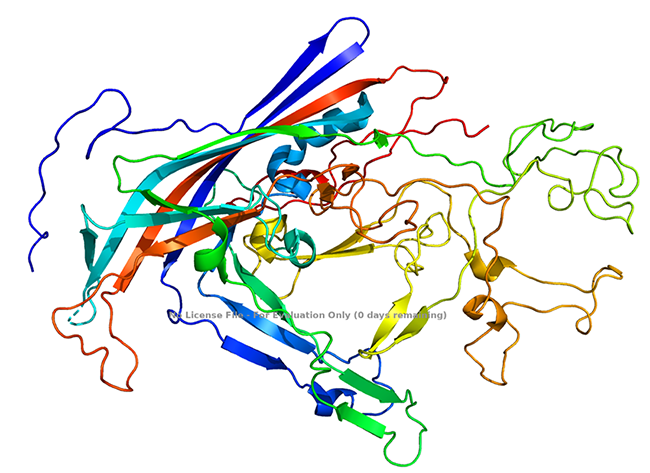
Figure 2 Sturcture of Canine Parvovirus VP2 protein (Xie and Chapman, 1996)
Notably, VP1 encompasses the complete sequence of VP2, along with an additional 143 amino acids at its N-terminus. A small number, typically five or six copies, of these N-terminal sequences of VP1 are protected from antibody binding, and their exact placement within the T1 icosahedral structure remains unidentified. In DNA-containing capsids, certain VP2 proteins can undergo proteolytic cleavage, resulting in the formation of a ~63-kDa protein known as VP3, which involves the removal of approximately 19 amino acids from the N-terminus.
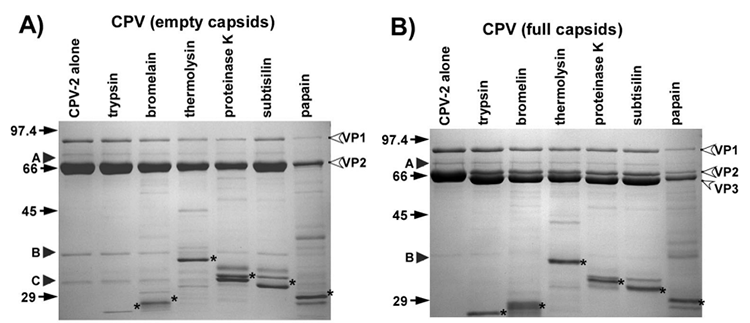
Figure 3 Sensitivity of the CPV-2 capsids to digestion with the proteases (Nelson et al., 2008)
The principal cell receptor for feline panleukopenia virus (FPV) and canine parvovirus (CPV) is the host's transferrin receptor (TfR). Both FPV and CPV capsids can attach to the feline TfR. However, CPV capsids have the additional ability to bind to the canine TfR, which plays a crucial role in determining the virus's ability to infect canine hosts. The interaction with the canine TfR is influenced by specific amino acid residues located at several distinct positions on the capsid surface, notably at positions 93, 299, and 323 of the VP2 protein.
Nonstructural protein-1 (P12929)
The NS1 protein of Canine Parvovirus Type 2 (CPV-2) is a multi-functional phosphoprotein that located at nucleus of the host cell. This protein exhibits both endonuclease and helicase activities, is essential for several critical processes, including the initiation and regulation of viral DNA replication, packaging of viral DNA, and the activation of viral promoters.
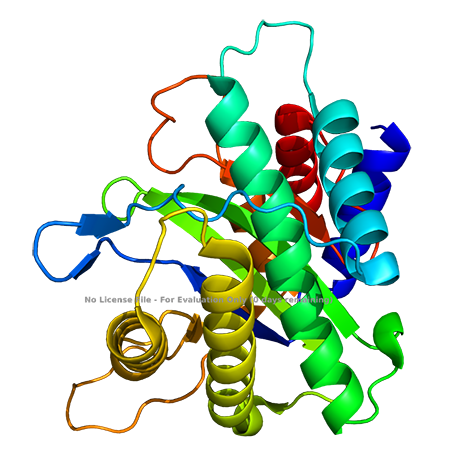
Figure 3 Crystal structure of CPV-2 NS1 homologous protein (Tewary et al., 2015)
Nonstructural protein-2
The nonstructural protein-2 (NS2) of canine parvovirus (CPV) is produced from the left-hand open reading frame of the viral genome and contains 87 amino-terminal amino acids in common with nonstructural protein 1 (NS1) joined to 78 amino acids from an alternative open reading frame. NS2 of minute virus of mice plays a role in controlling capsid protein assembly and translation in a host-specific manner. However, very little is known about the function of CPV-2 NS2 protein.
Reference
1. Callaway, H.M., Subramanian, S., Urbina, C.A., Barnard, K.N., Dick, R.A., Bator, C.M., Hafenstein, S.L., Gifford, R.J., and Parrish, C.R. (2019). Examination and Reconstruction of Three Ancient Endogenous Parvovirus Capsid Protein Gene Remnants Found in Rodent Genomes. Journal of virology 93.
2. Decaro, N., and Buonavoglia, C. (2012). Canine parvovirus--a review of epidemiological and diagnostic aspects, with emphasis on type 2c. Vet Microbiol 155, 1-12.
3. Govindasamy, L., Hueffer, K., Parrish, C.R., and Agbandje-McKenna, M. (2003). Structures of host range-controlling regions of the capsids of canine and feline parvoviruses and mutants. Journal of virology 77, 12211-12221.
4. Guerra, P., Valbuena, A., Querol-Audi, J., Silva, C., Castellanos, M., Rodriguez-Huete, A., Garriga, D., Mateu, M.G., and Verdaguer, N. (2017). Structural basis for biologically relevant mechanical stiffening of a virus capsid by cavity-creating or spacefilling mutations. Scientific reports 7, 4101.
5. Halder, S., Nam, H.J., Govindasamy, L., Vogel, M., Dinsart, C., Salome, N., McKenna, R., and Agbandje-McKenna, M. (2013). Structural characterization of H-1 parvovirus: comparison of infectious virions to empty capsids. Journal of virology 87, 5128-5140.
6. Hartmann, S.R., Charnesky, A.J., Fruh, S.P., Lopez-Astacio, R.A., Weichert, W.S., DiNunno, N., Cho, S.H., Bator, C.M., Parrish, C.R., and Hafenstein, S.L. (2023). Cryo EM structures map a post vaccination polyclonal antibody response to canine parvovirus. Communications biology 6, 955.
7. Kontou, M., Govindasamy, L., Nam, H.J., Bryant, N., Llamas-Saiz, A.L., Foces-Foces, C., Hernando, E., Rubio, M.P., McKenna, R., Almendral, J.M., et al. (2005). Structural determinants of tissue tropism and in vivo pathogenicity for the parvovirus minute virus of mice. Journal of virology 79, 10931-10943.
8. Lee, H., Callaway, H.M., Cifuente, J.O., Bator, C.M., Parrish, C.R., and Hafenstein, S.L. (2019). Transferrin receptor binds virus capsid with dynamic motion. Proceedings of the National Academy of Sciences of the United States of America 116, 20462-20471.
9. Mietzsch, M., Penzes, J.J., and Agbandje-McKenna, M. (2019). Twenty-Five Years of Structural Parvovirology. Viruses 11, 362.
10. Mira, F., Purpari, G., Lorusso, E., Di Bella, S., Gucciardi, F., Desario, C., Macaluso, G., Decaro, N., and Guercio, A. (2018). Introduction of Asian canine parvovirus in Europe through dog importation. Transboundary and emerging diseases 65, 16-21.
11. Nandi, S., and Kumar, M. (2010). Canine parvovirus: current perspective. Indian journal of virology : an official organ of Indian Virological Society 21, 31-44.
12. Nelson, C.D., Minkkinen, E., Bergkvist, M., Hoelzer, K., Fisher, M., Bothner, B., and Parrish, C.R. (2008). Detecting small changes and additional peptides in the canine parvovirus capsid structure. Journal of virology 82, 10397-10407.
13. Organtini, L.J., Allison, A.B., Lukk, T., Parrish, C.R., and Hafenstein, S. (2015). Global displacement of canine parvovirus by a host-adapted variant: structural comparison between pandemic viruses with distinct host ranges. Journal of virology 89, 1909-1912.
14. Tewary, S.K., Liang, L., Lin, Z., Lynn, A., Cotmore, S.F., Tattersall, P., Zhao, H., and Tang, L. (2015). Structures of minute virus of mice replication initiator protein N-terminal domain: Insights into DNA nicking and origin binding. Virology 476, 61-71.
15. Tsao, J., Chapman, M.S., Agbandje, M., Keller, W., Smith, K., Wu, H., Luo, M., Smith, T.J., Rossmann, M.G., Compans, R.W., et al. (1991). The three-dimensional structure of canine parvovirus and its functional implications. Science 251, 1456-1464.
Xie, Q., and Chapman, M.S. (1996). Canine parvovirus capsid structure, analyzed at 2.9 A resolution. Journal of molecular biology 264, 497-520.

Applications: ELISA, Immunogen, SDS-PAGE, WB, Bioactivity testing in progress
Expression system: E. coli
Accession: P17455
Protein length: Phe188-Tyr727

Host species: Mouse
Isotype: IgG2a
Applications: ELISA
Accession: P17455,P04864
Host species: Mouse
Isotype: IgG2a
Applications: ELISA
Accession: P17455,P04864
Host species: Dog
Isotype: IgG2, kappa
Applications: Blocking, ELISA
Accession: P61826
Host species: Caninized
Isotype: IgG2, kappa
Applications: Research Grade Biosimilar
Expression system: Mammalian Cells