Venezuelan, Eastern, and Western equine encephalitis viruses (VEEV, EEEV, and WEEV) are mosquito-borne zoonotic alphaviruses responsible for causing severe encephalitis in both equids and humans. These enveloped, positive-sense single-stranded RNA viruses are transmitted through mosquito vectors during their enzootic cycles, with reservoir hosts typically including rodents and birds. Despite their significant clinical impact, which can result in severe disease and mortality, there are currently no approved therapeutics available for the treatment of these infections. Unlike other alphaviruses, which are associated with arthritic syndromes, equine encephalitis viruses are primarily known for inducing severe encephalitic manifestations in humans.
Western equine encephalitis virus (WEEV) was first identified in equines in 1930 and reported in humans in 1937. Venezuelan equine encephalitis virus (VEEV) was first detected in equines in 1936, with human cases reported in the 1960s. Eastern equine encephalitis virus (EEEV) disease was first discovered in 1831 in Massachusetts, while the virus was first isolated from an infected horse’s brain in 1933. The mortality rates for VEEV, EEEV, and WEEV are <1%, 30-70%, and 3-7%, respectively.
The receptor for Eastern equine encephalitis virus (EEEV) is the very low-density lipoprotein receptor (VLDLR), and apolipoprotein E receptor 2 (ApoER2) also facilitates EEEV entry. In contrast, Venezuelan equine encephalitis virus (VEEV) utilizes the low-density lipoprotein receptor class A domain containing 3 (LDLRAD3) as its receptor. The receptor for Western equine encephalitis virus (WEEV) has been newly identified as protocadherin-10 (PCDH10).
The RNA genomes of these viruses are approximately 12 kb in length and encode two open reading frames. The first open reading frame encodes four nonstructural proteins (nsP1, nsP2, nsP3, and nsP4), while the second encodes five structural proteins (Capsid, E1, E2, E3, and 6K).
Fig.1 Equine encephalitis virus (EEV) particle structure and genome organization. (Han et al., 2023)
mRNA-capping enzyme nsP1
NsP1 is a cytoplasmic enzyme that facilitates mRNA capping by adding a 7-methyl-GMP cap to the mRNA and is involved in initiating minus-strand RNA synthesis. It also serves as a membrane anchor for the replication complex and induces filopodia formation by remodeling the host cell cytoskeleton. Palmitoylated nsP1 localizes to the host cell's filopodia and cytoplasmic vesicles, which may be linked to the formation of endosomal/lysosomal spherules for replication.
Protease nsP2
Nsp2 consists of two subdomains: an N-terminal helicase subdomain and a C-terminal protease subdomain. The C-terminal protease specifically cleaves polyproteins, releasing mature proteins. It is critical for viral replication and propagation, making it an attractive target for drug discovery.
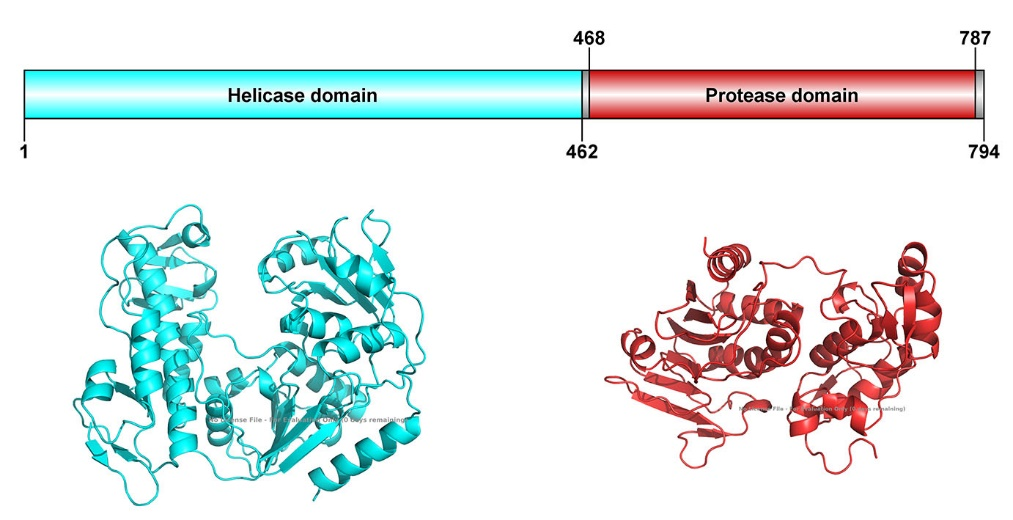
Fig. 2 Domain organization of nsP2 and structures predicted by Alphafold
Non-structural protein 3
NsP3 is comprised of three domains: an N-terminal macro domain (also called X domain), an alphavirus unique domain, and a C-terminal hypervariable domain (HVD). The macro domain is also found in SARS-CoV and SARS-CoV-2 nsp3. The HVD of nsP3 is intrinsically disordered, hypervariable, and hyper-phosphorylated on serine, threonine, and tyrosine residues. The HVD is critical for its replication in mosquito cells.

Fig. 3 Domain organization of nsP3
RNA-directed RNA polymerase nsP4
RNA-dependent RNA polymerase replicates both genomic and antigenomic RNA by recognizing specific replication signals. The early replication complex, formed by the polyprotein P123 and nsP4, synthesizes minus-strand RNAs. In contrast, the late replication complex, composed of fully processed nsP1 to nsP4, is responsible for producing genomic and subgenomic plus-strand RNAs.
Capsid
The equine encephalitis virus (EEV) has a diameter of approximately 70 nm and exhibits T=4 icosahedral symmetry. The viral RNA is enclosed by 240 copies of the capsid protein, which plays a crucial role in binding the negatively charged RNA genome via its N-terminal domain (NTD). The C-terminal domain of the capsid interacts with surface glycoprotein. equine encephalitis virus (EEV) capsid also function as a protease, enabling its self-cleavage from the structural polypeptide during translation. Additionally, the capsid shuttles between the nucleus and cytoplasm and is susceptible to nuclear trafficking inhibitors, highlighting its potential as an antiviral target.
Glycoprotein E1
E1 is a Class II viral fusion protein involved in low pH-mediated fusion with the host membrane. In the mature virion, E1 remains inactive as long as it is bound to E2. After the virus attaches to the target cell and undergoes endocytosis, the acidification of the endosome triggers the dissociation of the E1/E2 heterodimer, leading to the trimerization of E1 subunits. This E1 trimer becomes fusion-active and facilitates the fusion of the viral and endosomal membranes, promoting the release of the viral nucleocapsid into the cytoplasm. Efficient fusion requires the presence of cholesterol and sphingolipid in the target membrane.
Glycoprotein E2
The E2 protein plays a key role in receptor binding and is essential for viral entry. It is highly glycosylated, and its glycan chains interact with host cell DC-SIGN and L-SIGN molecules via its basic amino acids, facilitating viral attachment. It is initially synthesized as a precursor, which is processed by furin at the cell membrane just before virion budding. After cleavage, E1 and E2 form a stable heterodimer complex. Upon exposure to low pH, the mature E2-E1 heterodimer dissociates.
Glycoprotein E3
The E3 protein associates with the E1–E2 spike under acidic conditions to stabilize the heterodimer and inhibit premature fusion. At neutral pH in the extracellular medium, furin-cleaved E3 dissociates from the E1–E2 heterodimer. E3 provides the signal sequence for the translocation of the E3/E2 precursor protein to the host endoplasmic reticulum. Furin-cleaved E3 remains associated with the E1 spike glycoprotein and protects it from premature pH-induced inactivation during transport through the secretory pathway. After virion release from the host cell, the assembly protein E3 is gradually released into the extracellular space.
Protein 6K
The 6K protein acts as a viroporin, participating in virus glycoprotein processing, cell permeabilization, and the budding of viral particles. It disrupts cellular calcium homeostasis, likely at the endoplasmic reticulum level, resulting in increased cytoplasmic calcium levels. Due to its lipophilic properties, the 6K protein is thought to influence lipid selection that interacts with the transmembrane domains of glycoproteins, affecting the bilayer's deformability required for the extreme curvature during budding. Although present in low amounts in virions, 6K constitutes about 3% of the viral glycoprotein content.
| Protein | |
| Catalog No. | Product Name |
| YVV34404 | Recombinant EEEV Spike glycoprotein E1 Protein, N-His |
| YVV34401 | Recombinant EEEV Spike glycoprotein E2 Protein, N-His |
| YVV34403 | Recombinant WEEV Spike glycoprotein E2 Protein, N-His |
| YVV34402 | Recombinant WEEV Spike glycoprotein E1 Protein, N-His |
| EVV34401 | Recombinant WEEV Spike glycoprotein E1 Protein, C-His |
| Antibody | |
| Catalog No. | Product Name |
| PVV34401 | Anti-EEEV Spike glycoprotein E2 Polyclonal Antibody |
| PVV34402 | Anti-WEEV Spike glycoprotein E1 Polyclonal Antibody |
| PVV34403 | Anti-WEEV Spike glycoprotein E1 Polyclonal Antibody |
References
- Foy, N.J., Akhrymuk, M., Shustov, A.V., Frolova, E.I., and Frolov, I. (2013). Hypervariable domain of nonstructural protein nsP3 of Venezuelan equine encephalitis virus determines cell-specific mode of virus replication. Journal of virology 87, 7569-7584.
- Han, L., Song, S., Feng, H., Ma, J., Wei, W., and Si, F. (2023). A roadmap for developing Venezuelan equine encephalitis virus (VEEV) vaccines: Lessons from the past, strategies for the future. Int J Biol Macromol 245, 125514.
- Hasan, S.S., Dey, D., Singh, S., and Martin, M. (2021). The Structural Biology of Eastern Equine Encephalitis Virus, an Emerging Viral Threat. Pathogens 10, 973.
- Kehn-Hall, K., and Bradfute, S.B. (2022). Understanding host responses to equine encephalitis virus infection: implications for therapeutic development. Expert Rev Anti Infect Ther 20, 1551-1566.
- Lundberg, L., Carey, B., and Kehn-Hall, K. (2017). Venezuelan Equine Encephalitis Virus Capsid—The Clever Caper. Viruses 9, 279.



