Marburg virus (MARV), also known as Lake Victoria marburgvirus, is one of the most lethal human pathogens, causing Marburg virus disease (MVD) in humans, which is frequently referred to in the literature as Marburg hemorrhagic fever (MHF). The largest outbreak was nearly 90% lethal. Due to the lack of approved vaccine or treatment, the World Health Organization (WHO) classifies the Marburg virus as a Risk Group 4 pathogen, necessitating containment within a biosafety level 4 (BSL-4) facility. The Centers for Disease Control and Prevention (CDC) categorizes it as a Category A Bioterrorism Agent.
Three primary MARV vaccine candidates-cAd3, MVA-BN-Filo, and MARV DNA-are currently undergoing phase 1 clinical trials. Various Marburg vaccine platforms, including Adenovirus, DNA, VLP, and rVSV, have shown protective efficacy in non-human primates (NHPs).
The main natural host of the Marburg virus (MARV) is Egyptian Rousette Bat (ERB). Marburg virus (MARV) is usually spread through direct contact with the body fluids of an infected person or animal. This can happen when fluids enter through broken skin, mucous membranes, or through injuries from needles or sharp objects contaminated with these fluids.
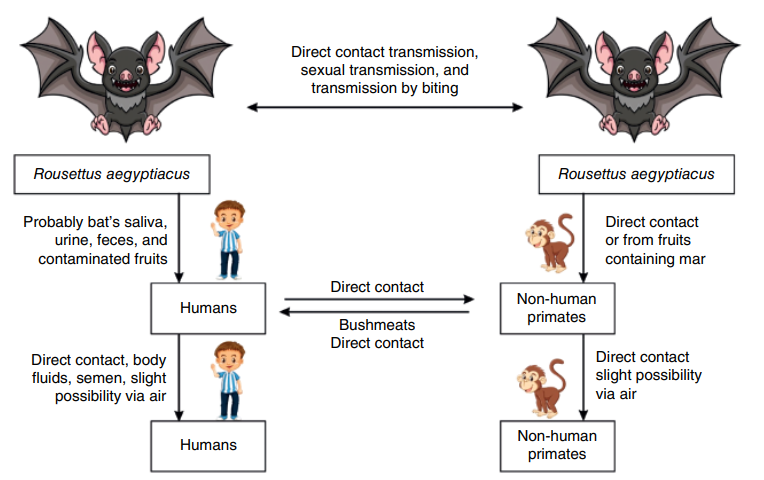
Figure 1. Cycle of transmission of the MARV between the bat host, monkey and humans. (Amponsah et al ., 2024)
Marburg virus (MARV) was identified by electron microscopy (EM) in 1967. The unusual filament-like structure caused some confusion, leading to speculation that it might be related to Leptospira, a spiral-shaped bacterium from the spirochaete family. Other scientists concluded that the pathogen was related to rhabdoviruses and named it Marburg virus.
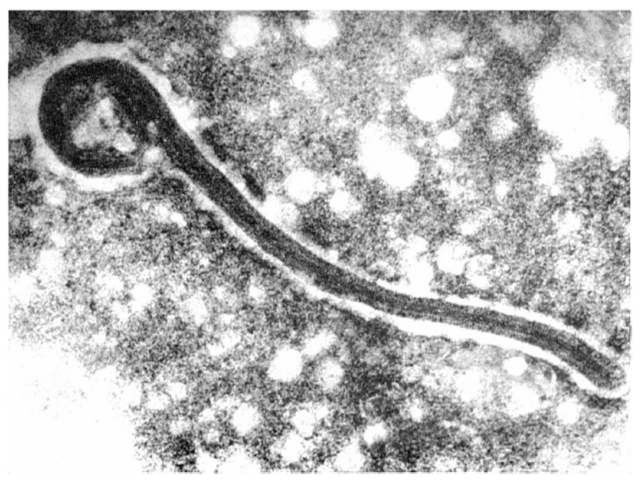
Figure 2. The first electron micrograph of a Marburg virus from 1967 (Brauburger et al., 2012)
MARV is an enveloped, non-segmented, single-stranded, negative-sense RNA virus. The length of MARV virions varies from 800 nm-1400 nm, have a variety of shapes, they might appear as U- or 6-shaped.The virion contains 7 proteins encoded by a 19 kb genome. The Marburg virus particles contain ribonucleoprotein (RNP) complexes composed of genomic RNA. Beneath the viral envelope, the virus includes structural proteins such as nucleoprotein (NP) and Nucleocapsid (VP35). Additional components are the transcription factor (VP30), nucleocapsid (VP24), and RNA-dependent RNA polymerase (L). The matrix protein (VP40) forms a regular layer, while glycoproteins (GP) create spikes on the surface of the virion. These spikes are about 7 nm in diameter and spaced approximately 10 nm apart.
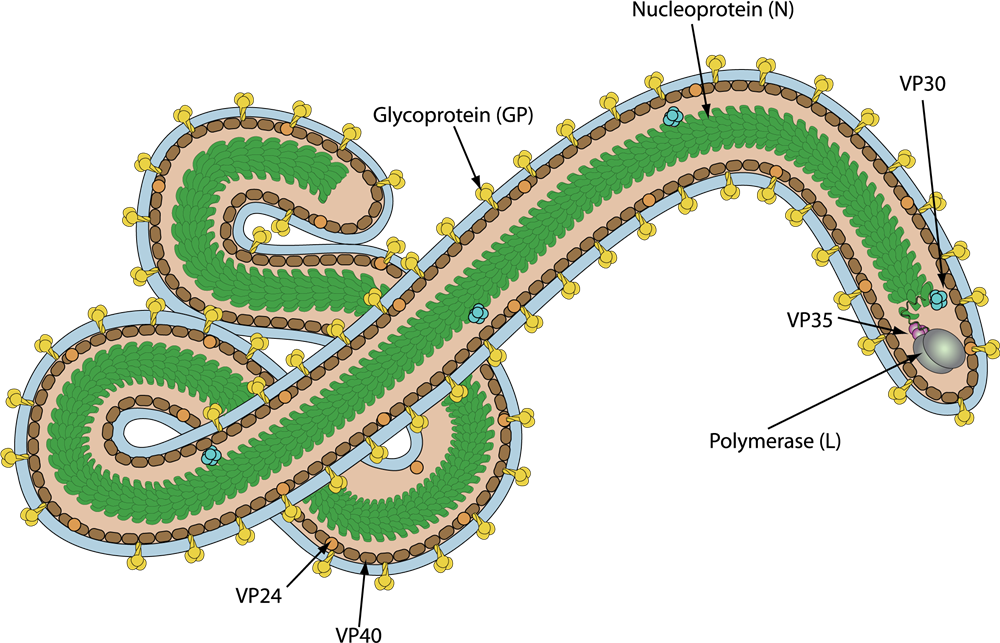
Figure 3. Schematic representation of Marburg virus (viralzone)
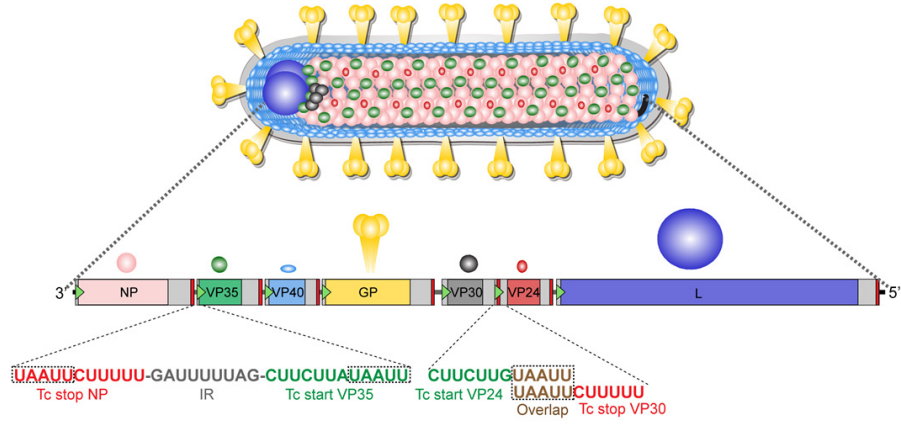
Figure 4. Marburg virion structure and genome organization (Brauburger et al., 2012)
Nucleoprotein (NP)
The nucleoprotein (NP) of MARV, the most abundant protein, consisting of 695 amino acids, encapsidates the vRNA to form a left-handed helical complex. This helical NP-RNA complex acts as a scaffold for the formation of the nucleocapsid, which is responsible for the transcription and replication of the vRNA. NP can interact with the VP35, VP40, VP30, and VP24 proteins. It can self-assemble into helical structures and is essential for nucleocapsid formation. NP is heavily phosphorylated at serine and threonine residues clustered in seven regions in the C-terminal part of the protein. Only the phosphorylated form of NP is incorporated into virions.
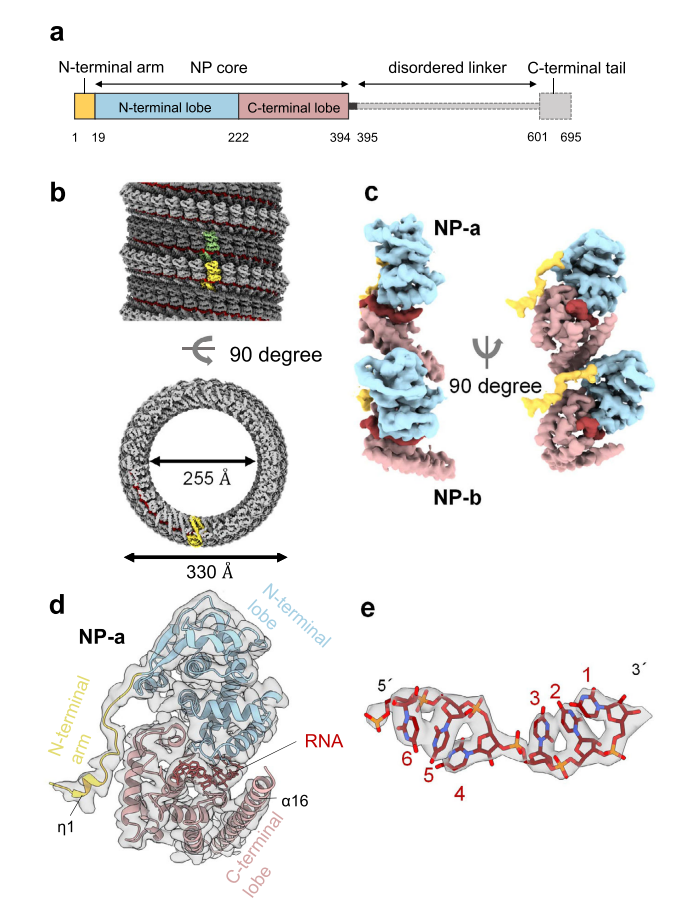
Figure 5. cryo-EM structure and atomic model of MARV nucleoprotein (1–395) (Fujita-Fujiharu et al., 2022)
Nucleocapsid (VP35)
VP35 is a polymerase cofactor and essential for transcription and replication. VP35 forms homo-oligomers mediated by a coiled-coil motif. It can bind to both NP and L. The C-terminal dsRNA-binding domain (RBD) of VP35 also acts as an IFN antagonist and also plays a role in immune evasion. Expression of MARV VP35 in the absence of other viral proteins is sufficient to completely block the induction of IFNα in stimulated human dendritic cells.

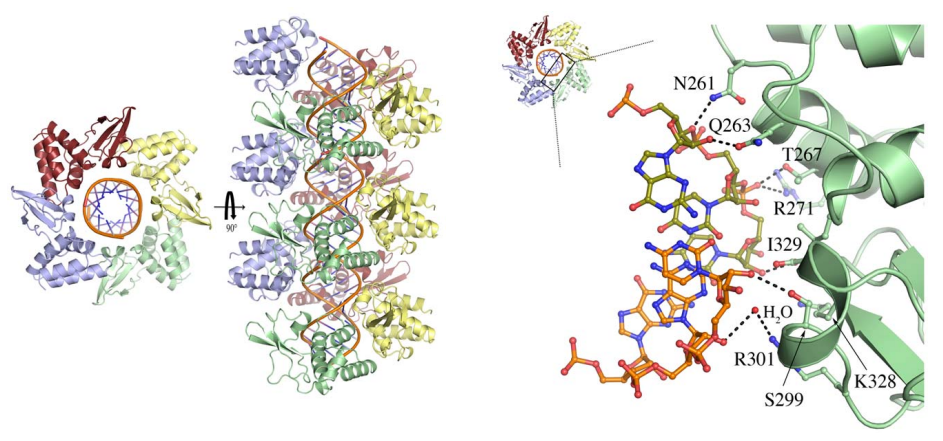
Figure 6. Domain organization and cyrstal structure of MARV VP35 (Bale et al., 2012)
Matrix (VP40)
MARV matrix protein VP40 is essential for assembly and release of newly replicated viruses and also suppresses immune signaling in the infected cell. MARV VP40 forms a dimer in solution, mediated by N-terminal domains, and that the formation of this dimer is essential for budding of virus-like particles. The N-terminal domain is necessary and sufficient for immune antagonism. The C-terminal domain of MARV VP40 are dispensable for immunosuppression but are required for virus assembly. The C-terminal domains are only 16% identical to those of Ebola virus, differ in structure from those of Ebola virus, and form a distinct broad and flat cationic surface that likely interacts with the cell membrane during virus assembly. VP40 is the Marburg virus matrix protein, which builds the protein shell beneath the viral envelope and confers the hallmark filamentous morphology to the Marburg virion. VP40 alone is able to induce the assembly and budding of filamentous viruslike particles (VLPs), which resemble authentic virions
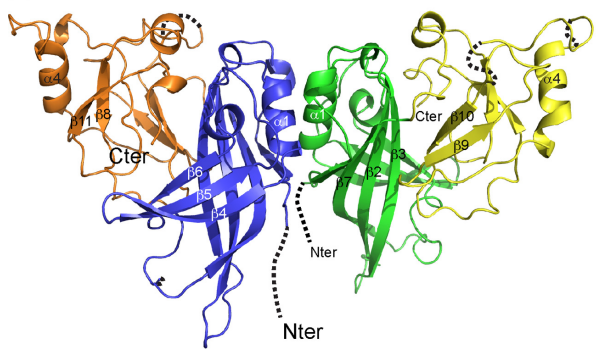
Figure 7. Crystal structure of Marburg virus VP40 ((Oda et al., 2016))
Glycoprotein (GP1 and GP2)
The glycoprotein (GP) is responsible for the attachment to and entry into target cells, making it the primary target for antibodies and vaccines. GP is a Type I transmembrane protein which is inserted into the viral envelope as homotrimeric spikes.
The GP precursor is cleaved by furin in the trans Golgi network resulting in two disulfide bond linked subunits: GP1 and GP2. The larger GP1 subunit contains the receptor-binding core, a "glycan cap" subdomain, and a C-terminal, heavily glycosylated mucin-like domain. GP2 contains the fusion peptideand is responsible for viral fusion.
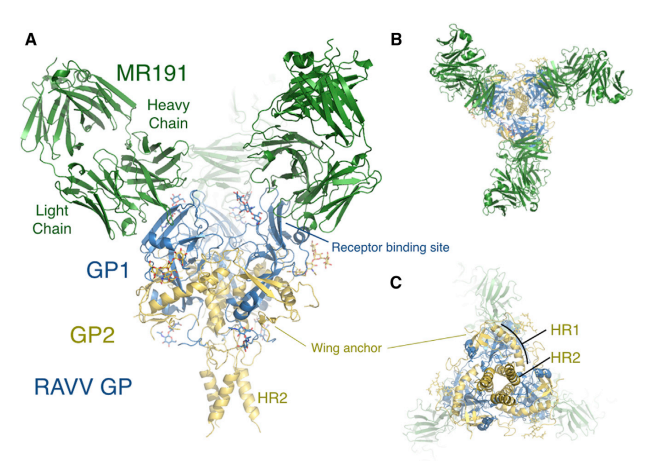
Figure 8. Neutralization antibody MR191 binds to MARV glycoprotein (King et al., 2018)
Transcription factor (VP30)
VP30 is tightly associated with the nucleocapsid through its binding to NP. It is highly phosphorylated at N-terminal serine and threonine residues, and this phosphorylation is essential for its interaction with NP. VP30 contains a C3H1 zinc finger domain, which is crucial for its role as a transcription initiation factor.
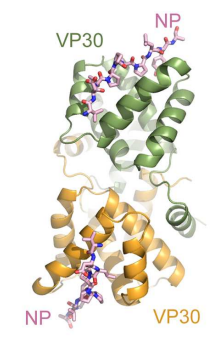
Figure 9. MARV VP30 CTD forms a dimer of globular domains and binds MARV NP peptide in the same location as EBOV(Kirchdoerfer et al., 2016)
nucleocapsid (VP24)
Unique among other Filoviridae viruses, VP24 of Marburg virus(MARV) is not immunosuppressive. It is generally considered as a minor matrix protein. But cryo-EM analysis of virus particles showed that VP24 is located close to nucleocapsid complex. VP24 is important for the release of viral particles during infection.
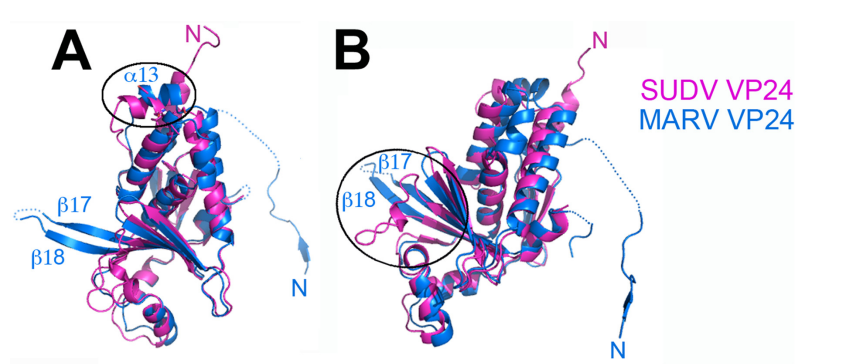
Figure 10. Differences between Marburg virus and ebolavirus VP24s (Zhang et al., 2014)
RNA-dependent RNA polymerase (L)
Protein L is the major component of the MARV polymerase complex. It functions as an mRNA capping enzyme and contains the polymerase catalytic domain. Together with VP35, Protein L forms the RNA-dependent RNA polymerase complex.
| Protein | |
| Catalog No. | Product Name |
| YVV24601 | Recombinant MARV VP40/Marburg VP40 Protein, N-His |
| YVV24501 | Recombinant MARV VP35/Marburg VP35 Protein, N-His |
| YVV24701 | Recombinant MARV VP30/Minor nucleoprotein VP30 Protein, N-His |
| YVV24801 | Recombinant MARV VP24/Marburg VP24 Protein, N-His |
| YVV24401 | Recombinant MARV NP/Nucleoprotein Protein, N-His |
| YVV24302 | Recombinant MARV GP2/Envelope glycoprotein 2 Protein, N-His |
| EVV24303 | Recombinant MARV GP2/Envelope glycoprotein 2 Protein, C-His |
| YVV24301 | Recombinant MARV GP1/Envelope glycoprotein 1 Protein, N-His |
| EVV24302 | Recombinant MARV GP1/Envelope glycoprotein 1 Protein, C-His |
| EVV24301 | Recombinant MARV GP/Envelope glycoprotein Protein, C-His |
| Antibody | |
| Catalog No. | Product Name |
| PVV24601 | Anti-MARV VP40/Marburg VP40 Polyclonal Antibody |
| RVV24501 | Anti-MARV VP35/Marburg VP35 Antibody (SAA1444) |
| RVV24401 | Anti-MARV NP/Nucleoprotein Antibody (SAA1443) |
| PVV24301 | Anti-MARV GP1/Envelope glycoprotein 1 Polyclonal Antibody |
| RVV24302 | Anti-MARV GP/Envelope glycoprotein Antibody (MR78) |
| RVV24301 | Anti-MARV GP/Envelope glycoprotein Antibody (MR191) |
Reference
1.Bale, S., Julien, J.P., Bornholdt, Z.A., Kimberlin, C.R., Halfmann, P., Zandonatti, M.A., Kunert, J., Kroon, G.J., Kawaoka, Y., MacRae, I.J., et al. (2012). Marburg virus VP35 can both fully coat the backbone and cap the ends of dsRNA for interferon antagonism. PLoS Pathog 8, e1002916.
2.Brauburger, K., Hume, A.J., Muhlberger, E., and Olejnik, J. (2012). Forty-five years of Marburg virus research. Viruses 4, 1878-1927.
3.Fujita-Fujiharu, Y., Sugita, Y., Takamatsu, Y., Houri, K., Igarashi, M., Muramoto, Y., Nakano, M., Tsunoda, Y., Taniguchi, I., Becker, S., et al. (2022). Structural insight into Marburg virus nucleoprotein–RNA complex formation. Nature Communications 13.
4.King, L.B., Fusco, M.L., Flyak, A.I., Ilinykh, P.A., Huang, K., Gunn, B., Kirchdoerfer, R.N., Hastie, K.M., Sangha, A.K., Meiler, J., et al. (2018). The Marburgvirus-Neutralizing Human Monoclonal Antibody MR191 Targets a Conserved Site to Block Virus Receptor Binding. Cell Host Microbe 23, 101-109 e104.
5.Kirchdoerfer, R.N., Moyer, C.L., Abelson, D.M., and Saphire, E.O. (2016). The Ebola Virus VP30-NP Interaction Is a Regulator of Viral RNA Synthesis. PLoS Pathog 12, e1005937.
6.Kortepeter, M.G., Dierberg, K., Shenoy, E.S., Cieslak, T.J., Medical Countermeasures Working Group of the National Ebola, T., and Education Center's Special Pathogens Research, N. (2020). Marburg virus disease: A summary for clinicians. Int J Infect Dis 99, 233-242.
7.Oda, S., Noda, T., Wijesinghe, K.J., Halfmann, P., Bornholdt, Z.A., Abelson, D.M., Armbrust, T., Stahelin, R.V., Kawaoka, Y., and Saphire, E.O. (2016). Crystal Structure of Marburg Virus VP40 Reveals a Broad, Basic Patch for Matrix Assembly and a Requirement of the N-Terminal Domain for Immunosuppression. J Virol 90, 1839-1848.
8.Ramanan, P., Edwards, M.R., Shabman, R.S., Leung, D.W., Endlich-Frazier, A.C., Borek, D.M., Otwinowski, Z., Liu, G., Huh, J., Basler, C.F., et al. (2012). Structural basis for Marburg virus VP35-mediated immune evasion mechanisms. Proc Natl Acad Sci U S A 109, 20661-20666.
9.Sharma, G., Sharma, A.R., and Kim, J.-C. (2024). Recent Advancements in the Therapeutic Development for Marburg Virus: Updates on Clinical Trials. Current Infectious Disease Reports 26, 57-67.
10.Zhang, A.P.P., Bornholdt, Z.A., Abelson, D.M., and Saphire, E.O. (2014). Crystal Structure of Marburg Virus VP24. Journal of Virology 88, 5859-5863.
All products are for research use only



