Background
Human T-cell leukemia virus type 1 (HTLV-1), like human immunodeficiency virus type 1 (HIV-1), belongs to the Retroviridae family. It is the first identified human oncogenic retrovirus and shares similar genomic structure and target cells (CD4⁺ T cells) with HIV-1. However, it exhibits strikingly different infection kinetics. HTLV-1 does not rely on persistent viral replication to sustain infection; instead, it achieves long-term persistence by "hijacking host cell proliferation mechanisms" and may ultimately induce adult T-cell leukemia/lymphoma (ATL). HTLV-1 is endemic in Japan, the Caribbean, Central Africa, and South America. For decades, the molecular mechanism underlying HTLV-1's spontaneous establishment of reversible latency has remained elusive.

Fig.1 HIV-1 and HTLV-1 virion composition(Viruses. 2022 Jan 14;14(1):152.)
A team from Kumamoto University, Japan, published groundbreaking research in Nature Microbiology titled "Intragenic viral silencer element regulates HTLV-1 latency via RUNX complex recruitment", revealing the key molecular switch for HTLV-1's establishment of long-term latent infection! They identified an open chromatin region (OCR) within the HTLV-1 provirus. This OCR recruits the host transcription factor complex RUNX1-CBFβ and corepressors (HDAC3/Sin3A), forming a transcriptional silencing complex that actively suppresses the activity of the 5'-LTR promoter, thereby maintaining latency. This study aimed to elucidate the core regulatory mechanism of HTLV-1 latency and explore the evolutionary basis for the differences in infection strategies between HTLV-1 and HIV-1.

Research Content
1. Discovery of a Novel Silencer Element: The OCR
Using ATAC-seq analysis on peripheral blood mononuclear cells (PBMCs) from HTLV-1-infected individuals and ATL patient samples, the research team identified an open chromatin region within the HTLV-1 proviral pol gene for the first time. Luciferase reporter assays confirmed that the OCR specifically suppresses the promoter activity of the 5'-long terminal repeat (5'-LTR) while having no significant effect on 3'-LTR-driven antisense transcription. This indicates that the OCR likely plays a key transcriptional repressive role in HTLV-1's unique latency regulation and low replication efficiency! Notably, no structurally conserved intragenic silencer was found in HIV-1 or other deltaretroviruses (e.g., HTLV-2, BLV).
2. Host RUNX Complex Mediates Silencing Mechanism
Combining chromatin immunoprecipitation sequencing (ChIP-seq) with motif analysis revealed the central role of transcription factor families RUNX1, ETS1, and GATA3 binding to the OCR, suggesting this mechanism may be the core molecular switch for HTLV-1's lifelong latency:
·The RUNX1-CBFβ complex acts as the key effector, directly binding to 3 conserved sites within the OCR via its Runt domain.
·Corepressors HDAC3/Sin3A are recruited to the OCR, mediating histone deacetylation and chromatin compaction.
·ETS1 acts as a molecular "switch" by antagonizing RUNX1 function; its downregulation is associated with transcriptional bursts.
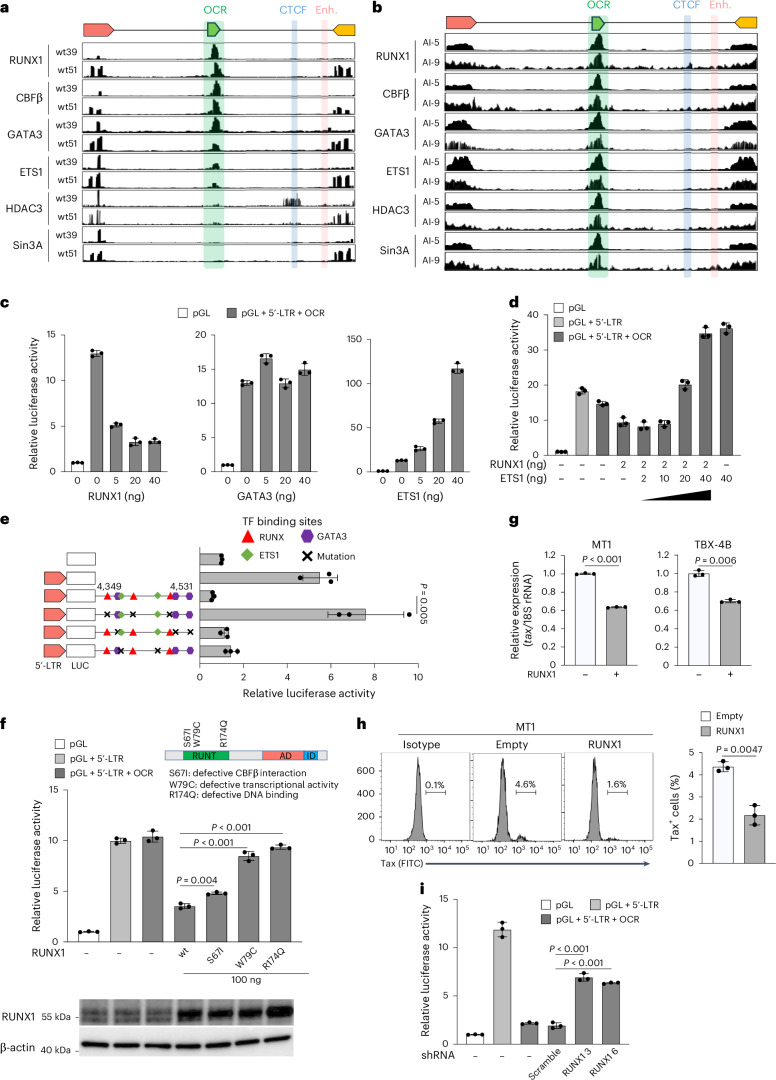
Fig.2 Molecular characterization of the silencer complex on the OCR
3. Validation of OCR's Role in Regulating Viral Latency
After constructing an HTLV-1 recombinant virus with mutated RUNX binding sites (s-mut), the following was observed:
·Tax mRNA expression increased by 52-fold, and viral particle production increased by 8-fold.
·Viral load continuously increased during long-term culture of the mutant virus, while the wild-type (WT) load decreased due to latency maintenance.
·Single-cell multiomics revealed that cells with an intact OCR formed a "latent cluster," while burst-state cells exhibited genome-wide chromatin openness.
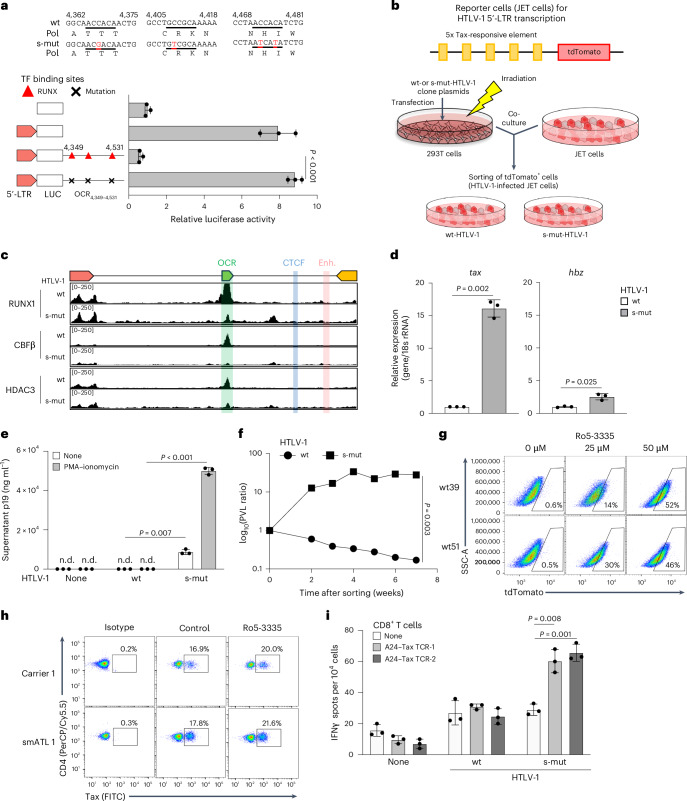
Fig. 3 Virological and immunological significance of the OCR function in HTLV-1 infection
4. Immune Evasion and Therapeutic Intervention Potential
OCR silencing significantly reduces viral antigen exposure:
·Treatment with the RUNX1 inhibitor Ro5-3335 increased the Tax-positive rate in latent cells by 46-fold.
·Cells infected with the OCR mutant virus were cleared by cytotoxic T lymphocytes (CTLs) with 20-fold higher efficiency.
·Cross-virus experiments confirmed the OCR's universality: Insertion into the HIV-1 nef gene region reduced HIV-1 p24 expression by 80% and abolished cytopathic effects, demonstrating that this element can reconfigure retroviral replication dynamics.
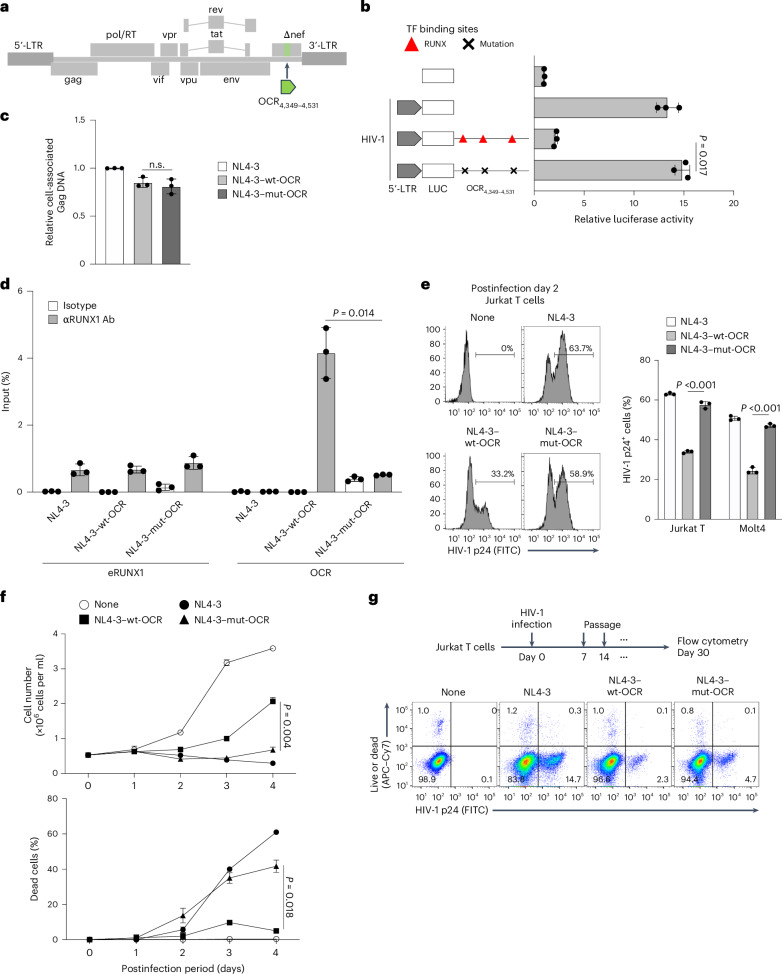
Fig. 4 Introduction of HTLV-1 OCR into rHIV-1 decreases proviral expression, virus production and cytopathic effect
Conclusions and Significance
This study is the first to elucidate the core mechanism by which HTLV-1 actively hijacks the host RUNX1-CBFβ-HDAC3/Sin3A complex via an "intragenic silencer" (OCR) to establish reversible latency:
Theoretical Level: Explains HTLV-1's unique strategy of relying on infected cell clonal expansion (rather than viral replication) for persistent infection, providing a new paradigm for virus-host co-evolution research.
Therapeutic Level: Targeting the OCR-RUNX1 interaction can specifically activate the latent viral reservoir, offering a novel target for combined immune clearance therapies.
Technical Level: The successful transplantation of the HTLV-1 silencer element into HIV-1 lays the foundation for engineering artificial latency regulation tools.
This study fills a critical knowledge gap in the latency mechanisms of exogenous retroviruses and has landmark significance for ATL prevention strategies and the development of latency reversal therapies!
References
- Kalinichenko S, Komkov D, Mazurov D. HIV-1 and HTLV-1 Transmission Modes: Mechanisms and Importance for Virus Spread. Viruses. 2022;14(1):152.
- Kenji Sugata, Rahman A, Koki Niimura, et al. Intragenic viral silencer element regulates HTLV-1 latency via RUNX complex recruitment. Nature Microbiology. Published online May 13, 2025.
AntibodySystem provides HTLV1&HIV-1-related proteins and antibodies, delivering more tools and solutions for mechanisms of exogenous retroviruses research.
HTLV1-Recomibinant Protein
|
Catalog |
Product Name |
|
YVV45201 |
Recombinant HTLV1 Capsid protein p24/p24 Protein, N-His |
|
YVV45302 |
Recombinant HTLV-1 Glycoprotein 46/gp46 Protein, N-His |
|
YVV45303 |
Recombinant HTLV-1 Glycoprotein 46/gp46 Protein, N-His |
HTLV1-Antibody
|
Catalog |
Product Name |
|
PVV45301 |
Anti-HTLV-1 Glycoprotein 46/gp46 Polyclonal Antibody |
|
MVV45301 |
Anti-HTLV-1 Glycoprotein 46/gp46 Monoclonal Antibody (m1A05) |
|
RVV45301 |
Anti-HTLV-1 Glycoprotein 46/gp46 Antibody (SAb2285) |
|
RVV45302 |
Anti-HTLV-1 Glycoprotein 46/gp46 Antibody (SAb2286) |
HIV-1 related prodcuts
|
Catalog |
Product Name |
|
YVV03201 |
Recombinant HIV-2 env/Env polyprotein/gp36 Protein, N-His |
|
EVV07801 |
Recombinant HIV1 gp120/SU Protein, C-His |
|
EVV03101 |
Recombinant HIV1 GP140 Protein, C-His |
|
RVV03101 |
Anti-HIV1 Surface protein gp120 Nanobody (SAA0912) |
|
DVV07801 |
Research Grade Anti-HIV gp120SU (VRC01) |
|
DVV03108 |
Research Grade Anti-HIV1 gp120/Glycoprotein 120 & CD4 Antibody (GS-8588) |
|
DVV03106 |
Research Grade Anti-HIV1 gp120/Glycoprotein 120 (VRC01LS) |
|
VVV19301 |
InVivoMAb Anti-HIV-1 Protease Antibody (F11.2.32) |
|
VVV03103 |
InVivoMAb Anti-HIV-1 gp140/Env Trimer Broadly Neutralizing Antibody (Iv0111) |



